Lo studio “Anthropogenic dispersal explains the phylogeography of insular edible dormouse Glis glis in the Mediterranean basin”, pubblicato su Mammal Research da Emiliano Mori, Leonardo Ancillotto, Andrea Viviano, Giovanni Amori, Fabrizio Vella e Flavio Monti dell’ Istituto di Ricerca sugli Ecosistemi Terrestri del Consiglio Nazionale delle Ricerche (IRET CNR), Davide Sogliani dell’università di Pavia, Giovanni Boano del Museo Civico di Storia Naturale di Carmagnola e Sandro Bertolino dell’università di Torino, parte da una constatazione: «La storia della colonizzazione umana nel Mediterraneo è stata a lungo riconosciuta come un fattore cruciale che influenza i modelli di biodiversità nell’Europa meridionale. Tuttavia, la nostra comprensione di come gli eventi di dispersione antropogenici e naturali abbiano interagito nel modellare la distribuzione della fauna selvatica, in particolare nei piccoli mammiferi, rimane limitata».
L’areale del ghiro (Glis glis), che si estende anche a diverse isole del Mediterraneo, fornisce l'opportunità di analizzare queste interazioni e il nuovo studio ha utilizzato il ghiro per testare ipotesi riguardanti i collegamenti tra la dispersione naturale e quella antropica a fini alimentari per modellare la distribuzione delle specie negli arcipelaghi del Mediterraneo.
Il ghiro Glis glis è un roditore di piccole e medie dimensioni (lunghezza del corpo 160–190 mm) e, nonostante preferisca gli habitat forestali è diffuso in un’are che va dalla Spagna settentrionale alla. I dati paleontologici indicano aree rifugio durante la glaciazione nella regione del Mediterraneo, con basso polimorfismo del gene del mtDNA a livello europeo probabilmente a causa della recente espansione, probabilmente avvenuta da un singolo refugium. In passato, la specie è stata introdotta intenzionalmente in varie località, comprese le isole, come risorsa alimentare e nell'Inghilterra centro-meridionale, come introduzione non intenzionale mediata dall'uomo.
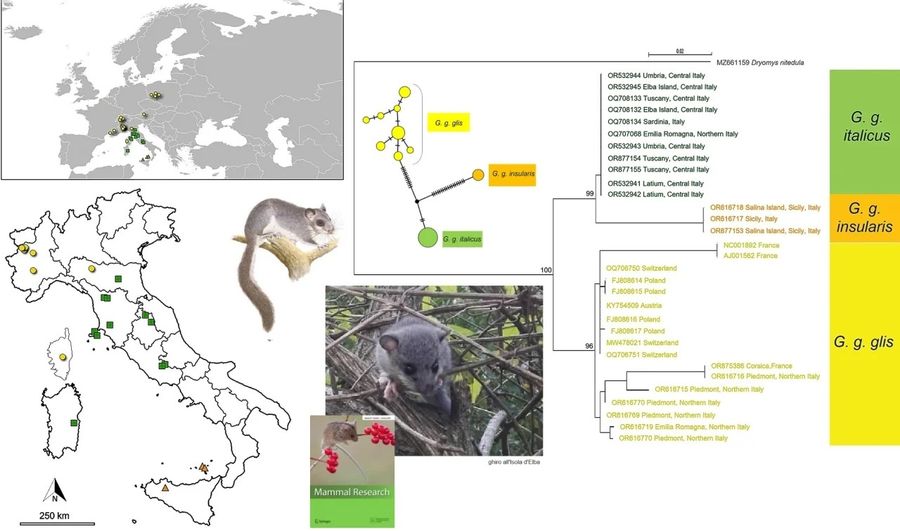
Sono state identificate 21 sottospecie di ghiro, ma le analisi filogenetiche e le caratteristiche morfologiche sembrano confermarne solo 5: G. g. glis , presente in gran parte dell'Europa; G.g. minutus , endemico della Macedonia del Nord; G.g. pindicus, confermato nell'isola di Alonissos ma forse presente anche nella Grecia continentale; G.g. italicus, endemico dell'Italia centrale e meridionale; G.g. insularis, endemico della Sicilia. Pertanto, sono stati descritti due cladi per le isole: G. g. insularis e G. g. pindicus.
In Italia vive il maggior numero di sottospecie di ghiro di tutta Europa: la sottospecie nominale si trova nelle regioni più settentrionali, la sottospecie italiana è presente nelle regioni centrali e meridionali e il clade siciliano è esclusivo della Sicilia. Lo studio “Italian Peninsula preserves an evolutionary lineage of the fat dormouse Glis glis L. (Rodentia: Gliridae)” pubblicato nel 2011 sul Biological Journal of the Linnean Society da Sabrina Lo Brutto, Maurizio Sarà e Marco Arculeolo dell’università di Palermo evidenziava che era necessario un ulteriore campionamento da popolazioni insulari (e/o geni aggiuntivi) della specie per chiarire le loro affinità filogenetiche con quelle continentali.
Il ghiro è infatti, è presente non solo in Sicilia e Sardegna ma anche in isole minori come Salina nell'arcipelago delle Eolie e l'Elba nell'Arcipelago Toscano, ma anche in Corsica. Sorprendentemente, lo studio “Diversity of small mammal communities of the Tuscan Archipelago: testing the effects of island size, distance from mainland and human density”, pubblicato nel 2015 su Folia Zoologica da Giovanni Amori (CNR), Vincenzo Rizzo Pinna (Systema Naturae), Giampiero Sammuri (Federparchi) e luca Luiselli (Demetra), rivelava il rinvenimento di resti di ghiro anche in una borra di barbagianni (Tyto alba) raccolta sull'Isola di Capraia nell'Arcipelago Toscano, anche se il ghiro potrebbe essere stato predato nella vicina Corsica o all’Isola d'Elba o nel continente e rigurgitato a Capraia in un secondo momento. Invece, la presenza del ghiro a Capri si basa esclusivamente sulla segnalazione di una guida turistica locale), e non è mai stata confermata scientificamente. La distribuzione del ghiro in Sardegna un tempo comprendeva un territorio più ampio e l'isola dell'Asinara, mentre attualmente la specie è presente solo nell'isola principale, dove è localmente classificata come “In pericolo di estinzione”.
Gli autori del nuovo studio sottolineano che «In questo lavoro si è mirato a chiarire le affinità filogenetiche del ghiro tra le popolazioni insulari e peninsulari. Abbiamo utilizzato un approccio barcoding, con particolare riferimento all'Isola d'Elba e alla Corsica, dove i ghiri non sono mai stati campionati per analisi genetiche. I risultati della nostra ricerca contribuiranno inoltre a una migliore comprensione dell’origine delle popolazioni insulari e faranno luce su possibili eventi di introduzione che potrebbero essersi verificati in passato».
I ricercatori hanno raccolto 21 campioni di ghiro, 2 dei quali all'Elba, 2 a Salina, 1 in Sardegna (Urzulei, Nuoro), 1 in Corsica (Vizzavona) e uno dalla Sicilia (Ficuzza, Palermo). Tutti gli altri campioni provenivano dall'Italia peninsulare. I campioni provenienti dall'Isola d'Elba e dalla Sardegna sono risultati raggruppati con campioni peninsulari dell'Italia centrale, mentre un secondo cluster era composto da campioni di Salina e siciliani, in linea con la sottospecie G. g. italicus e G. g. insularis. Campioni provenienti dal Nord Italia, comprese sia le Alpi che gli Appennini settentrionali, sono risultati raggruppati con il campione della Corsica in un sottocluster del G. g. glis che comprende anche sequenze provenienti da Polonia, Svizzera, Austria e Francia continentale.
I ricercatori evidenziano che «La nostra dimensione limitata del campione era dovuta alla piccola dimensione della popolazione del ghiro nelle isole italiane e in Corsica, e alla conseguente scarsità di esemplari nelle collezioni museali. Tuttavia, le nostre analisi hanno confermato la presenza di tre popolazioni divergenti, una presente nelle regioni settentrionali, una nelle regioni peninsulari e in Sardegna e la terza in Sicilia».
I risultati dello studio sono in linea con i lignaggi genetici identificati nello studio “Mitochondrial phylogeography of the edible dormouse (Glis glis) in the western Palearctic region”, pubblicato nel 2010 sul Journal of Mammalogy da un team internazionale di ricercatori guidato da Helene Hürner dell’Institut de Botanique del Belgi, confermando la classificazione sottospecifica stabilita. L'aplotipo italiano settentrionale appartiene alla sottospecie nominale Glis glis diffusa in tutta Europa, sebbene costituisca un sottoclade separato e divergente rispetto al resto dei campioni europei, che comprende anche il campione corso. Nell'Italia centrale, compresa la Sardegna, l'unico aplotipo presente è il G. g. italico. In SIcilia e nell'Isola di Salina c’è un terzo aplotipo distinto, G. g. insularis, che rappresenta il taxon gemello di G. g. italico . Questi tre aplotipi mostrano una distribuzione geografica ben definita e non sovrapposta da nord a sud. Ma la bassa variabilità genetica dei COXI per l'Italia peninsulare (G. g. italicus) e la Sicilia (G. g. insularis) potrebbe essere dovuta ad un effetto del fondatore. E il team della Hürner ha riscontrato una parziale sovrapposizione in Sicilia tra G. g. italicus e G. g. insularis.
Il nuovo studio conferma la bassa differenza genetica tra i gruppi insularis e italicus e quindi l'affinità genetica tra questi gruppi evidenziata dall'albero filogenetico. Il che suggerisce che «Potrebbe esserci stato un flusso genetico dall'Italia continentale alla Sicilia, dove una diversa unità tassonomica si è recentemente differenziata. Inoltre, come osservato per diverse altre specie, la sottospecie nominale del ghiro in Italia formava un sottogruppo divergente rispetto ai campioni provenienti dal resto d'Europa, coerentemente con il ruolo di rifugio glaciale della penisola italiana». Per stimare in modo affidabile l'origine temporale dei tre gruppi bisognerebbe ampliare l’indagine genetica.
Precedenti studi filogenetici sul ghiro hanno indicato un polimorfismo genetico estremamente limitato, rendendo la specie potenzialmente molto vulnerabile ai futuri cambiamenti climatici che potrebbero rappresentare una minaccia per la sua conservazione a lungo termine. I ricercatori fanno notare che «In questo contesto, l’Italia assume un ruolo fondamentale per la conservazione e il potenziale evolutivo del ghiro, poiché ospita la più grande diversità di aplogruppi a livello europeo».
La distanza genetica dell'aplotipo del gruppo meridionale rappresentato dai campioni siciliani è in linea con il ruolo riconosciuto svolto dalle regioni meridionali e dalle isole come rifugi glaciali. Evidenze paleontologiche suggeriscono che la dispersione dei rappresentanti della famiglia Gliridae sia avvenuta probabilmente solo nelle isole della parte occidentale del bacino del Mediterraneo, durante il Pleistocene superiore.
Per quanto riguarda gli esemplari provenienti dall’Isola d'Elba e dalla Corsica, i ricercatori scrivono che «I nostri risultati hanno rivelato che, nonostante la loro vicinanza geografica e distanza limitata, come nel caso della Sardegna, appartengono a due cluster diversi. I campioni dell'Isola d'Elba rientrano nel clade italicus , mentre il campione corso sembra appartenere alla sottospecie nominale distribuita nel resto d'Europa. Il campione corso mostra una maggiore somiglianza con gli esemplari dell'Italia settentrionale, in particolare del Piemonte, rispetto ai vicini campioni dell'Italia centrale e a quelli della Francia continentale. Questa dissomiglianza può essere indicativa di eventi di introduzione distinti sulle due isole in tempi storici. Nel caso della Corsica, l'introduzione è avvenuta probabilmente con individui originari del Nord Italia, e potrebbe essere avvenuta in tempi recenti, circa 100-150 anni fa, sebbene alcuni documenti storici risalgano anche a 4000-5000 anni fa. Tuttavia, sono necessari ulteriori campioni per confermare questa ipotesi».
Lo studio ricorda che «La prima segnalazione conosciuta di ghiro all'Isola d'Elba risale agli inizi del XIX secolo», ma durante le fasi finali dell'ultima era glaciale, molte isole vicine alla costa, come l'Elba, rimasero collegate alla terraferma per un lungo periodo. «Questa connettività avrebbe facilitato la diffusione delle specie continentali in queste isole – scrivono gli scienziati - il che potrebbe spiegare la somiglianza genetica osservata. Tuttavia, la mancanza locale di reperti fossili fa ipotizzare un'introduzione dalla costa dell'Italia centrale».
L'attuale distribuzione del ghiro è il risultato sia di processi di differenziazione evolutiva sia di eventi di introduzione mediati dall'uomo nei tempi antichi. I ricercatori spiegano che «Storicamente, il ghiro era comunemente importato e tenuto in grandi recinti murati noti come “glirarie” per l’ingrasso prima di essere consumato come prelibatezza. Questa pratica probabilmente ha facilitato la traslocazione di individui in varie località, tra cui la Sardegna e l'isola di Salina e l'emergere di popolazioni con struttura genetica distintiva in diverse aree. Il processo di introduzione potrebbe aver coinvolto individui provenienti da diverse regioni, spiegando potenzialmente la mescolanza di cladi, come G. g. italicus e G. g. insularis osservato in Sicilia». Nella maggior parte dei casi, le introduzioni recenti o antiche di ghiri nelle isole sono fallite. però lo studio evidenzia che «Tuttavia, in alcuni casi (Elba, Sardegna, Corsica) le attuali popolazioni di questo roditore sono geneticamente molto simili a quelle presenti sulla terraferma, indicando il successo delle introduzioni. Pertanto, se confermata anche attraverso geni nucleari, l'identificazione della sottospecie endemica sarda G. g. meloni potrebbe essere considerato un sinonimo junior di G. g. italicus. Diversamente dalla Sicilia, invece, l'isola di Salina sembra ospitare solo individui della sottospecie meridionale G. g. insularis, che potrebbe meritare particolari misure di conservazione».
I ricercatori concludono: «Gli esseri umani sono stati, e continuano ad essere, responsabili dell’alterazione dei modelli di dispersione naturale e del flusso genetico tra le popolazioni in molteplici modi. Il rimescolamento mediato dall’uomo negli areali di distribuzione delle specie e nell’integrità genetica è spesso inaspettato e difficile da monitorare, sottolineando l’importanza di una sorveglianza completa, integrativa in particolare per taxa poco studiati o trascurati come i piccoli roditori».
Umberto Mazzantini da greenreport.it